Le chariot
Le chariot

Serotonin
26 Dec 2016
Serotonin est l'amine biogenic qui est formée de l'acide aminé tryptophan par son hydroxylation et decarboxylation. Une quantité significative d'un serotonin contient dans les cellules enterokhromaffinnykh d'intestin, CNS, principalement dans hypothalamus et mesencephalon, thrombocytes, plus petite quantité dans le labrotsitakh, les cellules de mât, les glandes surrénales. Un serotonin exerce l'impact sur l'activité nerveuse, la réduction de causes de musculation lisse d'un intestin, un utérus, des bronches et aussi un rétrécissement de vaisseaux. Les réactions d'un organisme à un serotonin sont la pierre angulaire le central, myotropic, ganglionic, les effets réflexes.
Éducation. Un serotonin (5-gidroksi-triptamin, 5-HT) est synthétisé dans les cellules d'épithélium internes enterokhromaffinnykh d'un L-tryptophan. Un serotonin est aussi formé dans les cellules nerveuses de mezenterialny plexus et dans un CNS où les jeux un rôle de médiateur. Thrombocytes ne synthétisent pas un serotonin, le prennent cependant et accumulent.
Récepteurs de Serotonin. Il y a plusieurs types de récepteurs. Des récepteurs de point de vue pharmacologiques 5-HT1 et 5-NT2 et aussi 5-HT3 et 5-HT4 sont importants. En règle générale, les récepteurs travaillent par la G-protéine. Le récepteur 5-HT3 représente le canal cationic.
Effet de serotonin. Système cardiovasculaire. L'influence de serotonin sur le système cardiovasculaire est complexe puisque serotonin a l'effet (quelquefois en face renvoyé) dans beaucoup de sites par les types différents de récepteurs. Par 5-HT2-retseptory localisé dans les muscles non à raies de murs de vaisseaux, un serotonin provoque le rétrécissement de vaisseaux. L'effet en même temps indirect de serotonin peut mener à vasodilatation et à dépression de l'ABP : par 5-HT1A-retseptory il bloque des neurones sympathiques d'un brainstem et de la périphérie qui mène à la dépression de tonus sympathique ; l'activation de récepteurs 5-HT1 dans endothelium vasculaire mène à l'émission de vazodilatator (prostatsickin, NON). Le serotonin qui est libéré de thrombocytes joue un rôle dans les processus de thrombogenesis, hemostasis et dans hypertonia pathogenesis aux femmes enceintes.
Sumatriptan (un agent contre la migraine) est un 5-HT1D agonist et 5-HT1B-retseptorov. Le rétrécissement de vaisseaux crâniens provoqués par le blocus d'émission de neuropeptids mène à un soi-disant "neurogenic l'inflammation" ou diriger la crise vasculaire. En même temps il peut y avoir un sentiment de contrainte dans un sein attaché à un rétrécissement spasmodique d'artères coronaires. D'autres "tryptones" sont naratriptan, zolmitriptan et rizatripan.
Étendue digestive. Le serotonin formé dans mezenterialny plexus ou cellules enterokhromaffinnykh stimule un peristalsis et une sécrétion d'un intestin au moyen de l'influence sur 5-HT4-retseptory.
Les tentatives d'impact pharmacologique sur un motility d'un intestin par les récepteurs serotonin étaient inefficaces jusqu'à présent. Tsizaprid, un 5-HT4 agonist, cependant, peut augmenter motility propulsif d'un intestin. Cet effet secondaire est bloqué par Atropinum qui démontre le fait de monter de concentration d'Acetylcholinum sous l'influence de tsizaprid. Le médicament rend un ensemble d'effets secondaires. Il est inactivé sur le mécanisme avec la participation de SURZA4 qui mène aux actions réciproques imprévues différentes avec d'autres substances. Tsizaprid provoque des dérangements (en incluant sérieux) un rythme cordial (la réduction d'intervalles QT) donc le médicament n'a pas trouvé l'application.
Système nerveux central. Les récepteurs de Serotonin jouent un rôle important dans les fonctions différentes CNS.
Fluoxetine est l'antidépresseur bloquant le retour serotonin la reconsommation. Il augmente considérablement la motivation et baisse aussi l'appétit.
Sibutramin, blocker du retour neuronal serotonin reconsommation et Noradrenalinum ; il est appliqué comme le remède pour l'obésité.
Ondansetron a l'effet antiémétique exprimé au vomissement provoqué par l'application de tsitostatik. Il est un antagoniste de 5-HT3-retseptorov. Les analogues d'ondansetron sont tropisetron et granisetron.
L'acide et d'autres de psychedelia (psikhotomimetika), tel que la mescaline et psilocybin, provoquent des hallucinations, des désordres de conscience, peur, peut-être, à cause de l'activation de 5-NT-retseptorov.
Effets de serotonin
Le personnel d'université de McMaster est devenu des auteurs de travail scientifique pendant lequel le gène clé d'obésité - une sorte de serotonin a été trouvé. Il est célèbre que serotonin est "l'hormone de joie", son développement par un cerveau promeut la stabilité émotionnelle et la bonne humeur. Mais, comme les chercheurs canadiens expliquent, le serotonin qui est responsable des émotions plaisantes appartient au premier type de cette connexion.
"Il est divisé en deux types : dans la scène d'action et dans une forme de synthèse. Le premier type est développé dans un cerveau et influence de différentes émotions", - les biologistes ont expliqué.
Au deuxième type, serotonin périphérique appartient cette substance régule l'activité de tissu graisseux marron dont le développement d'obésité dépend. Vous pouvez aussi aimer Pankramin.
La graisse de Brown contient des composantes qui promeuvent la diminution dans le sucre dans le sang et aussi au brûlage de calories et à leur traitement dans l'énergie. Sur un corps de la personne il y a de certaines zones où la graisse marron s'arrange et ses cages sont plus actives, la figure à la personne est plus harmonieuse. Les chercheurs canadiens sont venus à la conclusion qu'en réprimant serotonin du deuxième type ; il est possible d'augmenter l'activité considérablement du métabolisme de cellules de tissu graisseux marron. Elle, à tour de rôle, forcera un corps "à brûler" de la graisse blanche � et, il se produira sans tenir compte comment la nourriture de haute calorie est mangée par la personne.
Récepteurs de Serotonin
Les effets de serotonin sont extrêmement différents. Cette substance sert d'un médiateur dans TsNS, les muscles lisses d'influences de vaisseaux et d'un CONARD, participe à trombotsitarnom vasculaire un homeostasis. Les méthodes pour le clonage moléculaire ont révélé l'à l'improviste grand nombre les récepteurs serotonin qui ont basé sur la structure et la fonction peut être divisée en 4 types. 5-T1--, 5-HT2-and 5-HT4-retseptory est intégré aux G-protéines et par ces protéines et les systèmes correspondants des deuxièmes intermédiaires influencent des fonctions d'enzymes différentes et des propriétés elek-trofiziologich de cages effector. Au contraire, 5-HT3-retseptory sont attachés aux canaux d'ion. Ici nous considérerons stimulators et blockers les récepteurs serotonin. Les plus nouvelles médecines de ces groupes qui font marcher sélectivement sur les sous-types séparés les récepteurs serotonin ont été reçues dans les travaux avec l'utilisation de récepteurs recombinant. Nous nous arrêterons aussi sur les modèles expérimentaux qui s'appliquent à une recherche des moyens influençant des fonctions mentales difficiles et leur persistance de violations, un comportement agressif, une inquiétude, une dépression, une vigilance de rêve de cycle et d'autre. stimulators sélectifs modernes de sous-types séparés les récepteurs serotonin déjà avec le succès sont appliqués en cas de la migraine et l'inquiétude et blockers sélectif en cas d'un certain nombre de violations gastrointestinal. Il est possible d'influencer des effets physiologiques de serotonin aussi au moyen des moyens opérant sur le transfert de serotoninergich. Donc les inhibiteurs de la capture de retour de serotonin étaient des médecines efficaces pour le traitement de dépression et d'inquiétude.
Malgré le fait que le rôle serotonin dans beaucoup de processus physiologiques et pathologiques ne lève pas des doutes, les points de son application et les mécanismes d'action sont étudiés mal. Peut-être, une telle situation est en partie provoquée par la variété le serotoninovykh de récepteurs. Ces récepteurs enregistrés au commencement par les méthodes pharmacologiques sont reçus par KDNK clonant aujourd'hui. Les récepteurs de Recombinant serotoninovy utilisent pour faire les études des mécanismes moléculaires d'effet de serotonin et aussi pour la recherche des agents qui influencent sélectivement des sous-types séparés de ces récepteurs. Le cercle d'une utilisation clinique d'agents semblables devient plus large et plus large.
Renseignements historiques
Dans 1930ème Erspamer a commencé à étudier la localisation de cellules enterokhromaffin au moyen des taches sur les dérivés indole. La plus haute concentration de tels dérivés a été enregistrée dans le muqueux par un CONARD ; plus loin il y avait thrombocytes et certains départements d'un CNS (Erspamer, 1966). Au bout d'un moment Peydzh et coauteurs, travaillant dans la clinique de Clevelend, ont émis pour la première fois la substance vasoconstrictive libérée de thrombocytes au cours d'un arrêt de saignement et ont déchiffré sa structure (Le rapport et autres, 1948). Cette substance appelée par Paige un serotonin (Page, 1976), a apparu à ces dérivés d'un indole qui a été enquêté par Erspamer. La description de voies de synthèse et de désintégration d'un serotonin (l'Uden-ami, 1959) et ses propriétés angiotonic (Sjoerdsma, 1959) a permis de faire une hypothèse selon laquelle les implications d'un soi-disant syndrome carcinoid aux patients avec les tumeurs des cellules enterokhromaffin sont provoquées par la production augmentée de cette substance. Vraiment, de tels patients ont une éjection quotidienne avec l'urine d'un serotonin et ses métabolites peuvent atteindre des centaines de milligrammes. Quelques symptômes de cette maladie indiquent dans une certaine mesure des mécanismes d'effet de serotonin. Donc aux patients la psychose semblable à l'acide survenant lors de la réception peut se développer. En estimant qui dans l'animal et les substances de tissus à légumes avec l'action hallucinogène sont trouvés semblables avec tripta-miny, il est possible de supposer que les substances semblables sont formées et provoquent une symptomatologie psychotique dans les patients avec le syndrome characinoid. De la fonction de médiateur d'un serotonin dans un cerveau de mammifères il a été suggéré au milieu les années cinquante (Brodie et Shore, 1957).
Les premières données sur les mécanismes moléculaires d'effet de serotonin ont été obtenues dans les expériences sur un foie de Fasciola hepatica (Mansour, 1979). Sous l'influence de serotonin à cela la mobilité et la concentration de tsAMF ont augmenté brusquement ; tant cela, qu'un autre effectue ont été bloqués par un acide. L'augmentation de la mobilité a été provoquée par phosphorylation tsAMF-dépendant de fosfofruktokinaza limitant glycolysis l'enzyme. Cependant les récepteurs serotoninovy négociant ces effets à un foie, probablement, d'autres, que les récepteurs de mammifères se sont connectés avec adenilattsiklaz à serotonin. Au plus tard ainsi les données exposées en détail sur les mécanismes d'effet de serotonin il n'a pas été succédé pour recevoir jusqu'à présent.
Serotonin a apparu aux usines et aux animaux déjà à de premiers stades d'évolution et à ceux-ci, peut-être, l'abondance de récepteurs serotonin parle (Peroutka et Howell, 1994). Le clonage de ces récepteurs a montré que quelques médecines qui ont été plus tôt considérées sélectives par rapport à leurs sous-types séparés ont en fait la haute affinité par rapport à plusieurs sous-types. Plus en détail de l'histoire d'étude et des effets de serotonin voir l'article Sjoerdsma et Palfreyman (1990).
Propriétés chimiques de serotonin
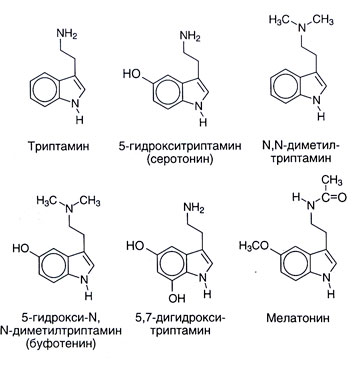
Sources. La structure chimique de serotonin et de quelques liens étroits à lui est donnée dans la fig. 11.1. Serotonin est répandu dans la vie d'animal et une usine : il est trouvé au vertébré, tunicates, les mollusques, arthropods, coelenterates, dans le fruit et dans les noix. C'est aussi trouvé dans les poisons � dans une ortie, dans les guêpes et les scorpions. De nombreux parents synthétiques ou naturels à la substance serotonin aussi dans un degré divers ont les effets physiologiques centraux et périphériques. Beaucoup Ni O-metilirovannye indolamines (par exemple, N, N-dimetiltriptamin) sont hallucinogens. Comme ils peuvent être développés dans un organisme, les considérait longtemps possibles responsable d'au moins quelques étalages de psychoses. Le mélatonine (5 metoksi N atsetiltriptamin) est formé de serotonin par N-acetylation avec l'O-methylation ultérieur (fig. 11.2). Cette substance sert d'indolamine principal de corps où sa synthèse est régulée par les facteurs externes (en particulier, le niveau d'illumination). Le mélatonine provoque depigmentation de melanotsit de peau et réprime la fonction d'ovaires. Peut-être, il joue un rôle dans les biorythmes et donc cela peut être utile à un syndrome de changement de fuseaux horaires.
Synthèse et catabolisme. Un serotonin est formé de l'acide aminé irremplaçable d'un tryptophan dans 2 stades (fig. 11.2). À la première étape sous l'influence de triptofangidroksilaza 5 hydroxytryptophan sont formés, c'est la limitation serotonin la réaction de synthèse. Triptofangidroksilaza représente un oxidase avec les fonctions mélangées. L'oxygène moléculaire et comme coenzyme tetrahydrobiopterine prend part à la réaction catalysée par cela. L'activité de triptofangidroksilaza, aussi bien que tyrosinehydroxylase, est régulée par phosphorylation, cependant le triptofangidroksilaza n'est pas inhibé par le produit fini sur le mécanisme de réaction négative. Dans un cerveau de triptofangidroksilaz il n'est pas saturé avec substrate et donc le taux de synthèse d'un serotonin dépend de la concentration d'un tryptophan. Le dernier vient aux cellules du cerveau par la capture active au moyen du transporteur qui est responsable du transport de plusieurs acides aminés neutres et branchus. À cet égard l'entretien d'un tryptophan dans un cerveau dépend pas seulement de sa concentration dans le plasma, mais aussi sur la concentration d'autres acides aminés rivalisant avec un tryptophan pour un transporteur.
Decarboxylation mène 5 hydroxytryptophans à la formation d'un serotonin. La longue dispute sur si decarboxylases 5 hydroxytryptophans et DOFA sont différents ou la même enzyme, a été résolue par KDNK le clonage des méthodes il s'est trouvé que le même produit de gène est responsable de decarboxylation de tous les deux substrates. Maintenant on appelle cette enzyme decarboxylase de L-acides-aminés aromatiques. C'est extrêmement répandu et affecte beaucoup de substrates. 5 hydroxytryptophan sont decarboxylized très vite et dans un cerveau il n'est pas presque trouvé. À cet égard les tentatives d'affecter la concentration dans un cerveau serotonin par la concentration changent 5 hydroxytryptophans sont condamnés à l'échec.
La voie principale de catabolisme serotonin la transformation dans l'acide 5-hydroxyindolacetic qui procède aussi dans 2 stades (fig. 11.2). Au début sous l'influence de MAO le 5-hydro-xyindolacetaldehyde qui passe alors dans l'acide 5-gidroksi-indolacetic sous l'influence d'enzyme aldegiddegidrogenaza répandue dans un organisme est formé (la quantité insignifiante 5 hydroxyindolacetaldehydes se transforment en alcool � 5-gidroksitriptofol). L'acide 5-hydroxyindolacetic est activly enlevé d'un cerveau ; ce processus est réprimé avec un inhibiteur nonspécifique de probenetsidy de transfert de transepithelial. Comme dans les cellules nerveuses presque 100 % de tous les métabolites d'une chute de serotonin à la part d'acide 5-hydroxyindolacetic, le taux d'un circuit d'un serotonin dans un cerveau est estimé lors du fait de monter de niveau d'acide 5-hydroxyindolacetic après l'introduction de probenetsid. L'acide 5-hydroxyindolacetic qui est formé dans un cerveau et d'autres organes et aussi des petites quantités d'un 5-gidroksitriptofol et de glucuronides est enlevé avec l'urine. Normal le quotidien egestion de l'acide 5-hydroxyindolacetic à l'adulte fait 2 � 10 mgs. Plus hautes valeurs � un signe fiable de syndrome carcinoid. La synthèse brusquement augmentée d'un serotonin à cette maladie demande de grands nombres de pyridinic nucleotides et un tryptophan et donc signes de manque d'acide nicotinic et d'un tryptophan pas une rareté à de tels patients. Les causes d'éthanol montant d'entretien de NADN et à la suite de 5 hydroxyindolacetaldehyde passent d'une voie s'oxydant d'un catabolisme à la récupération (fig. 11.2).
Ceux-ci sont lève egestion d'un 5-gidroksitriptofol un peu et réduit respectivement egestion d'acide 5-hydroxyindolacetic.
Il y a deux isoenzymes de MAO : MAO A et MAO B. Au début ils ont été divisés basés sur l'affinité aux substrats et la sensibilité aux inhibiteurs ; maintenant tant isoenzymes sont clonés, que les propriétés des formes clonées et naturelles étaient identiques (Shih, 1991 ; voir aussi hl. 10). MAO A a l'affinité préférentielle à serotonin et à noradrenaline et son inhibiteur sélectif est clorgilin. MAO B plus de travaux β-fenil-etilamin et benzylamine ; MAO B inhibiteur sélectif selegilin. L'affinité tant d'isoenzymes à la dopamine que de triptamin est identique. Les cages nerveuses contiennent tant MAO A que MAO B � principalement sur une membrane externe de mitochondrions. Puisque isoenzyme principal de plaquettes qui contiennent aussi serotonin dans la haute concentration sert MAO B.
Il était censé qu'il y a aussi d'autres voies d'un catabolisme de serotonin, par exemple sulphation et De - ou N-methylation. Le voyage final, pourrait mener en particulier, à la formation d'une substance psychotrope endogène 5-hydroxy-N, N-dimetiltriptamina (bufotenina, la fig. 11.1). Cependant d'autres metilirovanny indolamines (N, N-dimetiltriptamin, 5-metoksi-N, N-dimetiltriptamin) ont beaucoup plus exprimé des propriétés hallucinogènes et leur rôle dans pathogenesis de psychoses est plus probable.
L'inactivation de serotonin est exécuté pas seulement par la désintégration enzymatic, mais aussi au moyen de la capture de retour. Na + - le transporteur dépendant s'est positionné sur une surface externe de membrane presynaptic de la résiliation serotoninergich (fournit l'enlèvement de serotonin d'une fêlure synoptique) et une surface externe d'une membrane de plaquettes est responsable de cette capture (extrait serotonin du sang). Aux plaquettes c'est la voie unique d'action de remplissage de serotonin puisqu'il n'y a aucune enzyme de synthèse de cette substance dans eux. Le transporteur serotonin, aussi bien que d'autres transporteurs de monoamines, est cloné (hl. 12).
Points d'application serotonin
La plupart de partie de tout l'entretien de serotonin dans un organisme tombe à la part de tissus peripheric bien qu'il serve aussi comme un médiateur dans un CNS. Sa concentration dans les cellules enterokhromaffinnykh et thrombocytes est la plus haute. Un serotonin joue un rôle important dans un règlement d'un motility d'un CONARD.
Cellules d'Enterokhromaffinny. Ces cellules s'arrangent dans le muqueux un CONARD. Surtout il y a beaucoup d'entre eux dans un duodénum. Dans les cellules enterokhromaffinnykh il est synthétisé d'un tryptophan et un serotonin s'accumule et aussi d'autres agents biologiquement actifs, par exemple la substance P et kinina contiennent. Il y a un certain niveau de sécrétion basale d'un serotonin dans un CONARD. Cette sécrétion amplifie lors de l'étirement mécanique (par exemple, en entrant dans une nutrition ou une solution de sel hypertonique) et à une ennuyeuse de fibres de motif de nerfs vagus. Peut-être, l'effet stimulant de serotonin sur un motility d'un CONARD est aussi négocié par son influence sur les neurones de plexus intermusclé (Gershon, 1991 ; voir aussi hl. 38). La sécrétion brusquement augmentée d'un serotonin et d'autres agents biologiquement actifs au syndrome carcinoid est suivie par gastrointestinal correspondant, les dérangements cardiovasculaires et nerveux. En plus la synthèse augmentée d'un serotonin peut avoir pour résultat le manque d'acide nicotinic et d'un tryptophan.
Thrombocytes. Thrombocytes diffèrent d'autres éléments formulés d'un sang, en particulier, dans la capacité de prendre le magasin et libérer un serotonin. La synthèse d'un serotonin dans thrombocytes n'arrive pas. Un serotonin est pris thrombocytes d'un sang et vient pour le stockage au secret elektronop des grains au moyen du transport actif. Ces processus sont dans beaucoup de respects semblables à la capture et au stockage de Noradrenalinum dans les résiliations sympathiques (hl. 6 et 12). Par une membrane de thrombocytes un serotonin est transféré au moyen de Na + - le transport dépendant et aux grains � par le transport actif secondaire avec l'utilisation comme une source d'énergie d'un gradient électrochimique pour H + encadré par N +-Atfazoy. En même temps la concentration d'un serotonin dans les grains atteint 0.6 mol/l, c'est 1000 fois plus haut, que dans le cytoplasme de thrombocytes. Taux Sur + - reconsommation serotonin dépendante par thrombocytes un indicateur sensible d'activité d'inhibiteurs de reconsommation serotonin.
La fonction principale de thrombocytes est hemostasis : ils ferment des espaces dans endothelium endommagé. D'autre part, l'intégrité d'un endothelium joue un rôle important dans le fonctionnement de thrombocytes (Furchgott et Vanhoutte, 1989). L'endothelium contacte constamment à thrombocytes comme à cause des forces de changement opérant dans le sang actuel ils sont déplacés à la périphérie de vaisseaux (Gibbons et Dzau, 1994). L'effet de Vasoconstrictive de serotonin et tromboksan d'A2 sont opposés par un facteur endothelial de vasorelaxation (N0 et, peut-être, autres substances) (Furchgott et Vanhoutte, 1989 ; la fig. 11.4). Pour l'adhésion et l'agrégation de thrombocytes la condition d'un endothelium a l'importance cruciale (Hawiger, 1992 ; Ware et Heistad, L993). Quand thrombocytes sont contigus à endothelium endommagé, ils émettent les substances provoquant leur adhésion et libération d'un serotonin. Le RADIOGONIOMÈTRE AUTOMATIQUE et ò ð îìáîêñàí A2 appartiennent à de telles substances (hl. 26 et 55). Linkng d'un serotonin avec 5-HT2A-retseptorami a le faible effet proagregant qui amplifie brusquement en présence d'un collagen. Si le défaut d'un mur vasculaire atteint des couches de muscle lisses, donc un serotonin rend l'effet vasoconstrictive direct servant d'un de mécanismes hemostasis. Cet effet amplifie sous l'influence d'agents biologiquement actifs qui sont émis dans le domaine du dommage tromboksan d'A2, kinin, vasoactive peptides. La formation de thrombi à un atherosclerosis est promue par la destruction d'un endothelium et, par conséquent, le manque d'un facteur endothelial de vasorelaxation. Dans ces conditions la direction de processus à thrombogenesis procèdent de façon incontrôlée, comme un cercle vicieux. Un rôle dans eux est aussi joué par un serotonin. Le dessin semblable peut être observé à d'autres maladies de vaisseaux, par exemple le syndrome de Reynaud et vasospastic stenocardia.
La description à la fig. 11.4. Fonctions d'un serotonin de thrombocytes. La libération d'un serotonin de thrombocytes est commencée eux par l'adhésion et l'agrégation. À tour de rôle, un serotonin provoque 1) l'activation á-ÍÒ-receptors thrombocytes et, vrezultata, le changement d'une forme et l'accélération d'agrégation du dernier, 2) l'activation 5-HT, - les récepteurs semblables d'un endothelium avec l'allocation d'un facteur endothelial de vasorelaxation, 3) l'activation de S-HT-receptors de muscles non à raies de vaisseaux et de rétrécissement du dernier. Tous ces processus procèdent dans l'action réciproque avec beaucoup d'autres agents biologiquement actifs et mènent finalement au saignement de l'arrêt.
Système cardiovasculaire. La réaction typique de vaisseaux sanguins à serotonin se rétrécit. Les vaisseaux d'organes de CONARD, reins, poumons et cerveau sont surtout sensibles à cela. Serotonin provoque aussi la réduction de muscles non à raies de bronches. Ses effets sur le cœur sont différents que le changement de récepteurs serotonin de tonus de nerfs végétatifs et de réactions réflexes est expliqué par l'activation de différents sous-types (Saxena et Villalon, 1990). Donc chronotropic positif direct et effet inotropic de serotonin sur le cœur peuvent être déguisés par les effets d'exaltation des fibres allant des récepteurs de barre et de chemoceptors. L'influence d'un serotonin sur les résiliations afferent de nerfs vagus provoque l'Yarisha réflexe de Betsold que l'on montre bradycardia pointu et chute de l'ABP. Quelquefois arterioles sous l'influence d'un serotonin ne sont pas réduits et, au contraire, la suppression de libération de Noradrenalinum des résiliations sympathiques s'étendent à la suite de l'allocation d'un facteur endothelial de vasorelaxation et de Prostaglandinums et aussi. D'autre part, un serotonin renforce en soi l'action vasoconstrictive de Noradrenalinum, angiotensin 11 et Histaminum. Il promeut l'effet statique haemo encore plus efficace de serotonin (Gershon, 1991).
CONARD. Probablement, puisque la source principale et le stockage d'un serotonin dans un organisme servent des cellules enterokhromaffinny muqueuses un CONARD. Le serotonin alloué par ces cellules survit à une veine portique à un foie où il est transformé par métabolisme sous l'influence de MAO A (Gillis, 1985). Une quantité de serotonin passe un métabolisme hépatique, mais est vite prise un endothelium de capillaires pulmonaires et est aussi affectée par MAO. Le serotonin qui est alloué dans un mur d'organes d'un CONARD lors de leur étirement mécanique ou exaltation de nerfs vagus participe à un règlement local de ces organes. Sous l'influence d'un serotonin un motility d'un estomac et d'un intestin peut tant amplifier et être freiné (Dhasmana et autres, 1993) comme dans un CONARD il y a au moins 6 sous-types les récepteurs serotonin (l'étiquette. 11.2). L'effet stimulant de serotonin est provoqué par son action sur les résiliations des nerfs convenables aux couches musclées longitudinales et circulaires (5-HT4-retseptory), sur les neurones intra-muros (5-HTj-and 5-NT|R-retseptory) et est direct sur les muscles lisses (5-ÍÒ-receptors dans les intestins et 5-HT2B-retseptory dans le jour d'un estomac). Dans un gosier serotonin affecte 5-HT4-retseptory qui aux types différents d'animaux peut être suivi tant en réduisant, que la relaxation de muscles lisses. 5-HT3-retseptory (ce sont beaucoup de participants aux résiliations de fibres sensibles de l'errance et d'autres nerfs et aussi sur les cages) jouent un rôle clé dans un réflexe émétique (Grunberg et Hesketh, 1993). Dans une texture intermusclée les résiliations serotoninergetic sont trouvées. La libération de serotonin dans les intestins est provoquée par l'acétylcholine, l'irritation de nerfs sympathiques et l'augmentation de la pression intestinale intra et de la diminution dans le ph (Gershon, 1991). Le serotonin qui est émis en même temps, à tour de rôle, commence la réduction de peristaltic.
La région principale de concentration de corps de neurones serotonin dans CNS est des cœurs de couture de brainstem. Les processus de ces neurones vont à tous les départements d'une moelle épinière et de tête (hl. 12). Un serotonin est alloué pas seulement dans les résiliations presynaptic, mais aussi dans soi-disant varicosity d'axons où il n'y a aucun synapse exactement exprimé (Descarries et autres, 1990). Dans ces cas il affecte beaucoup de structures adjacentes immédiatement. Un tel trait d'allocation et un effet de serotonin seront constitués avec le point de vue répandu qu'un serotonin n'est pas seulement un médiateur, mais aussi le modulateur de neurone (hl. 12).
Dans les résiliations de neurones serotonin il y a toutes les composantes nécessaires pour la synthèse d'un serotonin d'un tryptophan (fig. 11.2). serotonin formé vient vite aux ampoules synoptiques où sur lui MAO ne peut pas travailler. Après la libération dans une fissure synoptique un serotonin est de nouveau pris la résiliation nerveuse au moyen de Na - un transporteur dépendant. Cette capture de retour sert d'un moyen efficace d'un inactivation d'un médiateur. Les mêmes molécules d'un serotonin qui ne reviennent pas à la résiliation nerveuse sont faites sauter par MAO localisé dans les neurones postsynaptic et les cellules suivantes.
Effets d'Electrophysiological. Ces effets d'un serotonin diffèrent par de différentes régions d'un cerveau et dans de différents neurones et dépendent sur quels récepteurs il affecte (l'étiquette. 11.3 ; Aghajanian, 1995). Un serotonin peut avoir l'excitant et l'effet de frein différant sur la dynamique temporaire sur les mêmes neurones. Donc dans les neurones hippocampus un serotonin provoque au début la polarisation hyper (provoqué par l'activation de 1A-récepteurs 5-HT), alors la dépolarisation lente (provoqué par l'activation de 5-HT4-retseptorov).
TsNS. Serotonin influence TsNS beaucoup de fonctions, en incluant un rêve, une activité cognitive, une perception, une direction de mouvements, contrôle thermal, sensibilité pénible, appétit, comportement sexuel et règlement endocrine. Dans un cerveau tous ont cloné des récepteurs serotonin sont trouvés et souvent au même département il y a plusieurs tels récepteurs. De plus, bien que l'expression de récepteurs serotonin dans les neurones séparés soit étudiée insuffisamment, il est possible de croire que sur le même neurone plusieurs sous-types de ces récepteurs peuvent être localisés et leur activation peut être suivie tant par synergistic que par effets opposés. Cela peut être la cause d'une variété extraordinaire d'influences de serotonin sur les fonctions du cerveau.
La polarisation de Hyper et la diminution dans la résistance d'une membrane survenant en cas de l'activation 5-HT) les A-récepteurs, sont provoqués par l'augmentation de la perméabilité de potassium. Ces effets sont bloqués par la toxine, mais ne dépendent pas de tsAMF. Donc, ils peuvent être provoqués par l'interface directe de 5-NT|A-retseptorov (par la G-protéine semblable à Gi) avec le canal de potassium (Andrade et autres, 1986). L'activation des récepteurs localisés sur un corps et des dendrites de neurones d'amandes d'une couture d'un tronc d'un cerveau mène aussi à K + - la polarisation hyper dépendante. Ici participe aussi à une émission j d'un signal d'un récepteur au canal sensible à la toxine de G-protéine, mais le courant de potassium a d'autres caractéristiques, que ce qui survient en cas de l'activation de postsynaptic 5-HT1A-retseptorov dans un hippocampus. Il n'est pas encore connu au moyen ce que l'activation de mécanismes 5-HT1D-ayTopeuerrropoB mène à la suppression de libération de serotonin. Probablement, le nombre des canaux calcic s'ouvrant en réponse à l'impulsion nerveuse les diminutions potentielles.
L'activation de 5-HT2A-retseptorov est suivie par la dépolarisation lente. Dans certains départements d'un cerveau (par exemple, dans le cortex préfrontal, une amande adjacente et une amande de motif d'un nerf du visage) cette dépolarisation est provoquée par la diminution dans la perméabilité de potassium (Aghajanian et autres, 1987). Il y a aussi d'autre mécanisme raccordé avec l'influence sur les canaux d'ion ; il mène à la croissance d'excitabilité de neurone et de renforcement de réaction aux médiateurs excitants (par exemple, un glutamate). Le rôle de système fosfoinozitid dans ceux-ci effectue n'est pas encore établi. Quand sur le même neurone il y a aussi 5-HT1-retseptory et 5-Nt2d-retseptory, la réaction finale à serotonin dépend d'un rapport entre la polarisation hyper provoquée par l'activation de 5-HT1-retseptorov et la dépolarisation provoquée par l'activation de 5-HT2A-retseptorov. Sur un fond de la polarisation 5-Nt2d-blokatorov hyper amplifie. Dans beaucoup de régions d'écorce 5-HT2A-retseptory sont localisés sur les neurones insérés de Gamkergichesky et sur les neurones pyramidaux. Donc, la stimulation de ces récepteurs peut mener aux influences multidirectionnelles sur les neurones pyramidaux dans la dépendance parce que si l'action sur ces neurones ou sur les neurones de Gamkergichesky prédominera. L'activation de 5-Nt2s-retseptorov sur oocytes de grenouille, l'express de m d'ARN de ces récepteurs, mène à la suppression de courant de potassium. Dans un cerveau un tel effet n'est pas encore révélé. La stimulation de 5-HT4-retseptorov qui est suivi par l'activation provoque aussi la dépolarisation lente provoquée par la diminution dans la perméabilité de potassium dans les neurones. Il n'est pas encore clair pourquoi deux types différents de récepteurs serotonin connectés aux systèmes différents des deuxièmes intermédiaires rendent le même effet physiologique. De plus, la dépolarisation lente est provoquée par l'activation d'encore un type de récepteurs serotonin 5 HT1P-récepteurs. Ces récepteurs sont accompagnés par le disponible seulement sur les neurones intestinaux intra et ont des propriétés pharmacologiques spéciales (Gershon, 1991).
L'activation de 5-HT3-retseptorov provoque la dépolarisation bystry. Il est provoqué en s'ouvrant du canal passant Na + et K + (Higashi et Nishi, 1982) et faisant un complexe uniforme (hemo le canal sensible) avec un récepteur. L'organisation semblable d'un semblable 5-HT3-retseptora à l'organisation N-holinoretseptora a été confirmée dans les expériences avec la fixation locale. 5-HT3-retseptory sont trouvés dans TsNS, ganglia sympathique, fibres afferent parasympathiques et sympathiques, intra les neurones intestinaux et les lignes cellulaires d'origine neyronal (par exemple, NG108-15). Sur les propriétés pharmacologiques 5-HT3-retseptory diffèrent d'autres des récepteurs serotonin ; peut-être, il y a plusieurs sous-types de ces récepteurs différant par les combinaisons différentes de subjunits.
Fonctions mentales. Les moyens opérant sur les récepteurs serotonin et les changements différents de fonctions mentales. Beaucoup de modèles expérimentaux destinés pour une estimation temporaire de la stimulation ou du blocage de l'activité de médecines par rapport à ceux-ci ou ces récepteurs, sont basés sur une recherche de tels actes de motif stereotypic comme, par exemple, le réflexe. Les techniques de Behaviouristic (par exemple, une méthode de choix de médecine) permettent de suggérer d'une composante subjective de réactions aux médicaments psychotropic. Ces techniques sont aussi utilisées pour une recherche des médecines opérant sur le transfert de serotonin, dans hallucinogens particulier (voir ci-dessous). L'analyse de nombre énorme des travaux consacrés à l'influence de serotonin sur le comportement est au-delà de notre livre et nous nous arrêterons seulement sur ces expériences qui ont un rapport direct sur psychopathology de la personne. Pour la connaissance plus détaillée de ce sujet il est possible de recommander Glennon aux révisions parfaites et Lucki (1988), Zifa et Fillion (1992), Koeketal. (1992).
Allez à vélo un rêve est la vigilance. Le règlement d'un cycle un rêve est la vigilance est devenu une des premières fonctions mentales pour lesquelles le rôle serotonin a été exactement établi. Après le travail classique sur les chats qui est réalisé par Mouret et ñîò ð. (Mouret et autres, 1967), l'ensemble des données que l'épuisement de stock serotonin au moyen des causes l'insomnie éliminée en entrant d'un prédécesseur de serotonin 5 d'apparus. Il a aussi semblé que tryptophan et pas stimulators sélectif de récepteurs serotonin raccourcissent le temps de somnolence et étendent la durée générale d'un rêve. Blockers de récepteurs serotonin peut tant monter et baisser une part d'un sommeil lent profond qui, probablement, est provoqué par l'action sur de différents sous-types de récepteurs serotonin (Wasquier et Dugovic, 1990). Tant aux animaux qu'à blockers 5-HT2A-and 5-Nt2s-retseptorov (par exemple, riganserin) provoquent plutôt de manière fiable l'augmentation d'une part d'un sommeil lent profond.
Agression et impulsivité. Les données obtenues tant sur les animaux que sur la personne, démontrez qu'un serotonin joue un rôle important dans le comportement agressif et impulsif. Dans beaucoup de communication d'essais cliniques entre le faible niveau de 5 - de l'acide hydroxyindolacetic dans SMZh et un tel comportement est montré (Brown et Linnoila, 1990). Donc la dépression de ce niveau est attachée aux tentatives impulsives de suicide (mais pas avec les pensées suicidaires ; Virkkunen et autres, 1995). Aussi bien qu'en cas de tous les autres effets d'un serotonin, les animaux n'ont aucune donnée finale sur l'influence d'un serotonin sur le comportement agressif encore bien que l'hypothèse d'une telle influence soit tout à fait prouvée. Récemment il y avait le soutien de données génétique et le fait de dilater de telles vues. Les récepteurs de Serotonin enquêtés par les méthodes pour le génie génétique 5-HT) le V-récepteur était le premier de. Par une recombinaison homologue la ligne de souris qui ont un gène codant ce récepteur a été reçue il a été inactivé (Saudau et autres, 1994). À de tels animaux l'agression la plus pointue s'est développée qui parle d'un rôle de ces récepteurs ou dans le fait de devenir les contours de neuronic qui sont responsables du comportement agressif ou c'est immédiat dans la plupart de tel comportement. À la personne la mutation pointée du gène codant MAO A est enregistrée ; l'agression en même temps extrême dans la combinaison avec le retard mental est aussi observée (Brunner etal., 1993). Comme cela a apparu, les implications correspondantes sont aussi disponibles pour les souris avec les mutations ayant pour résultat le manque de MAO A (Les cas et autres, 1995). Ces données, certainement, confirment une hypothèse d'un rôle de dérangements de transfert de serotoni-nergic dans le comportement agressif.
Inquiétude et dépression. Efficacité des moyens influençant serotoninergichesky transfert (par exemple, inhibiteurs de la capture de retour de serotonin), en cas de la frustration dérangeante et de la dépression l'évidence pour l'avantage d'un rôle serotonin dans pathogenesis de ces conditions. Cependant sur les modèles expérimentaux classiques de ceux-ci la frustration reçoit des résultats équivoques l'effet de médecine dépend des espèces et de la race d'un animal et de la technique appliquée. Donc agonist partiel de 5-HT1A-retseptorov buspiron (hl. 19), étant un tranquillisant efficace à la personne, ne réduit pas à l'inquiétude d'animaux dans les expériences livrées par une technique de préférence ou d'évitement ; pendant ce temps cette technique a été utilisée en cas du développement de tranquillisants. En même temps buspiron et d'autre blockers de 5-HT1A-retseptorov ont l'effet anxiolytic tout à fait efficace sur d'autres modèles de frustration dérangeante (Barrett et Vanover, 1993). Les travaux récents sur les souris avec les gènes inactivés de 5-NT1L-retseptorov sont aussi témoins pour l'avantage d'un rôle de ces récepteurs dans pathogenesis d'inquiétude et, peut-être, les dépressions (Les parcs et autres, 1998 ; Ramboz et autres, 1998). D'autre part, stimulators de certains les récepteurs serotonin, en incluant 5-T2A--, inquiétude de cause de 5-HT2C-and 5-HT3-retseptorov tant dans les animaux expérimentaux, que dans la personne. Le rôle de ces récepteurs et dans les modèles expérimentaux d'une dépression est censé (par exemple, l'impuissance formée).
La personne de données directes sur un rôle serotonin dans un pathogenesis d'une dépression n'a toujours pas assez. En même temps il y a des faits cliniques très brillants. Donc aux patients avec des effets de dépression d'inhibiteurs du retour serotonin la reconsommation sont éliminés vite aux influences réduisant serotonin le niveau dans un cerveau. En tant que tel le service d'influences, par exemple, la réception de parachlorphenylalanine ou des boissons qui ne contiennent pas un tryptophan, mais riche avec les acides aminés neutres (Delgado etal., 1990). Il est intéressant que ces influences ne provoquent pas en soi et n'aggravent pas une dépression. Cela signifie que le niveau suffisant d'un serotonin dans un cerveau est nécessaire tout d'abord pour l'efficacité d'inhibiteurs de sa capture de retour.
Agents influençant serotonin niveau dans les tissus
Dans les expériences renvoyées sur la clarification d'un rôle physiologique d'un serotonin il est possible d'appliquer blockers de récepteurs serotonin ou les agents influençant serotonin le niveau dans les tissus. Jusqu'à récemment dans la majorité de travaux seulement les médicaments du deuxième des groupes mentionnés étaient des mécanismes utilisés d'action de blockers les récepteurs serotonin ont été étudiés mal.
Lors de l'entretien bas d'un tryptophan dans un régime serotonin le niveau dans un cerveau diminue, au haut contenu d'augmentations de tryptophan. Comme la réaction de limitation de synthèse d'un serotonin est catalysée, les inhibiteurs de cette enzyme provoquent la chute pointue de niveau d'un serotonin. Utilisez le plus souvent un inhibiteur sélectif irréversible d'un triptofangidroksilaza parachlorphenylalanine. Sous l'influence de cette substance serotonin le niveau considérablement et diminue pendant longtemps et l'entretien de catecholamins ne change pas.
Parachloramphetamine et d'autres amphétamines halogenated provoquent l'émission de serotonin de thrombocytes et de neurones. Dans un cerveau après qui vient là la longue dépression de réserves de serotonin. Les amphétamines de Halogenated sont largement utilisées dans les travaux expérimentaux. Deux d'entre eux fenfluramin et decsfenfluramin ont été appliqués comme anorecsant, cependant dans la connexion avec les rapports sur leur action toxique cardio en 1998 ils aux Etats-Unis ont été retirés de la vente. Les conséquences d'utilisation de ces agents sont à la hauteur de la fin non étudiée. Dans les neurones du cerveau serotonin ils provoquent exprimé et long (jusqu'à plusieurs semaines) la dépression de niveau d'un serotonin et du contenu de protéines, spécifiques à ces neurones qu'un transporteur d'un serotonin et de triptofangidroksilaza diminue en même temps. Il peut démontrer l'action neurotoxin, cependant les signes de mort de neurones sous l'influence d'amphétamines halogenated ne sont pas trouvés. Dérivés de triptamin avec les députés supplémentaires dans l'Indo-lin un anneau (par exemple, 5.7-digidroksitriptamin ; voir la fig. 11.1) provoquent indubitablement la mort les neurones serotonin. L'introduction d'un 5.7-digidroksitriptamin un animal adulte est mené à la destruction sélective par les résiliations serotonin, cependant les corps de neurones ne souffre pas et au fil des années les résiliations régénèrent. Au contraire, aux animaux nouveau-nés tant les résiliations que les corps de neurones serotonin et donc neogenesis n'arrive pas périssent.
Les inhibiteurs du retour serotonin la reconsommation, par exemple fluoxetine appartiennent aux agents qui influencent précisément le transfert de serotonin. Le mécanisme de leur action se compose à l'extension d'effets du serotonin alloué lors de l'initiation des résiliations nerveuses. Si avec ces médicaments pour entrer dans 5 hydroxytryptophan, donc les influences de serotonin amplifient brusquement. Les inhibiteurs du retour serotonin la reconsommation sont un des antidépresseurs les plus modernes et répandus. L'inhibiteur du retour serotonin la reconsommation, Noradrenalinum et Dofaminum sibutramin est appliqué comme anoreksant. Dans un organisme deux métabolites actifs que, probablement et ont l'effet thérapeutique en sont formés. Il n'est pas encore clair, l'influence sur quel médiateur a provoqué l'effet de sibutramin.
Les inhibiteurs de MAO et de Reserpinum appartiennent à pas les agents sélectifs influençant serotonin le niveau dans les tissus. Les inhibiteurs de MAO bloquent la voie principale d'un métabolisme d'un serotonin et Reserpinum provoque son émission du dépôt de neurone avec l'usure ultérieure. Tous ces agents mènent à la dépression exprimée d'entretien de serotonin, cependant en même temps dans le même degré aussi le niveau de diminutions de catecholamine. Donc que les agents pour les inhibiteurs d'analyse pharmacologiques de MAO et de Reserpinum sont utilisés rarement. Ils ont été appliqués dans la psychiatrie : Reserpinum comme narcoleptic et inhibiteurs MAO comme les antidépresseurs.